Evidence Against Evolution
(c) Copyright 2000, David A. Duncan
Fossil data
Fossil Formation
The great majority of fossils are found in sedimentary rock. Sedimentary rock is formed from particles which are carried by water and deposited when the water action becomes sufficiently still for the particles to settle out. Once deposited, the action of chemical cementing agents and/or pressure causes the material to become rock. Plants and animals that are swept into this process can thus become fossilized as part of the rock. Sedimentary rock layers vary from a few feet to hundreds of feet thick and somewhat rarely as much as 1000 feet thick.
Fossil Dating
Geologists have observed that in sedimentary rock, that layers are readily identified that most often contain the same type of fossilized animal remains. Observing that this layering has the less complex organisms at the bottom and proceeds to more complex organisms at the top, scientists have theorized that this sedimentary layering provides primary evidence of a time sequence of when these organisms lived. By assuming that rock formation has always occurred at the same rate that it occurs today, scientists have devised a time scale for the various layers. According to this theory, the formation of sedimentary rock that is hundreds of feet thick would take millions of years. Due to this observation coupled with the advent of evolutionary theory that also requires millions of years, the estimated age of the earth by evolutionary geologists has increased greatly in the last 150 years. The application of radiometric dating methods has somewhat standardized the age estimates to about 4.5 billion years.
Geologists believe that certain fossils were formed during restricted time spans and therefore these “index fossils” are used as primary evidence to date rocks based on their position in the layer sequence. This arrangement of the layers into a standardized sequence is known as the “geological column.”
The Sequence in the Column
With fossils formed in sedimentary rock, the expectation of a world-wide deluge would be the formation of sediments across a large portion of the land mass. “About three-fourths, perhaps more, of the land area of the earth, 55 million square miles, has sedimentary rock as the bedrock at the surface or directly under the cover of mantle-rock …The vast bulk of the stratified rocks is composed of shallow-water deposits.”[1]
If the sequence found in the geological column is not produced by evolution, then what caused the sequence? Morris and Whitcomb, in “The Genesis Flood” provide three reasons for the sequence:
(1) Marine Life. The sea bottoms being affected by the “opening of the fountains of the deep” would be first affected, and would likely then be first deposited. “With reference to the Cambrian strata, supposedly the oldest fossiliferous strata: ‘At least 1500 species of invertebrates are known in the Cambrian, all marine, of which 60% are trilobites and 30% brachiopods.’[2] … It is not until the Permo-Carboniferous is reached, well up in the geologic column, that the first land animals are encountered.”[3]
(2) Sorting. They point out that the dynamics of moving water (hydrodynamics) will lead to sorting of material based on particle diameter, sphericity, and density. “Particles which are in motion will tend to settle out in proportion mainly to their specific gravity (density) and sphericity. It is significant that the organisms found in the lowest strata, such as the trilobites, brachiopods, etc., are very ‘streamlined’ and are quite dense. … on the average, the sorting action is quite efficient and would definitely have separated the shells and other fossils in just such fashion as they are found, with certain fossils predominant in certain horizons, the complexity of such ‘index fossils’ increasing with increasing elevation in the column, in at least a general way”[4]
(3) Mobility. Vertebrates would be expected to be higher in the column since they possess higher mobility. In general, the larger the animal, the more likely it would be to escape the flood waters. This is what is found in the column, as is evidenced by the proposed evolution of the horse from a small fox-like animal to the large horse of today. The simplest vertebrates (ostracoderms) are found in the Ordovician strata. “Fishes are found in profusion in the Devonian, often in great sedimentary ‘graveyards,’ indicating violent deposition, and often in fresh-water deposits. It is obvious that fish do not normally die and become fossilized in such conditions as these but usually are either destroyed by scavengers or float on the surface until decomposed. The whole aspect of the fossil fish beds bespeaks violent burial in rapidly moving deltaic sediments.”[5]
Thus, in summary, the expected results of a world-wide deluge should be deposition in order of increasing mobility (ability to escape inundation), of decreasing density and other hydrodynamic factors, and of increasing elevation of habitat. “The order is exactly what is to be expected in light of the Flood account and, therefore, gives further circumstantial evidence of the truthfulness of that account … And the fact that, although this order is generally to be expected, it is found to have many exceptions, both in terms of omissions and inversions, is also certainly to be expected in terms of Deluge events, but is extremely difficult to account for logically in terms of evolution and uniformity.”[6]
Table 1: Geological Column
|
Era |
Period |
Epoch |
106 years |
|
Cenozoic |
Quaternary |
Holocene |
0-0.1 |
|
Pleistocene |
0.1-1.6 |
||
|
Tertiary |
Pliocene |
1.6-6.3 |
|
|
Miocene |
6.3-23 |
||
|
Oligocene |
23-36.6 |
||
|
Eocene |
36.6-53 |
||
|
Paleocene |
53-66 |
||
|
Mesozoic |
Cretaceous |
66-135 |
|
|
Jurassic |
135-205 |
||
|
Triassic |
205-250 |
||
|
Paleozoic |
Permian |
250-290 |
|
|
Carboniferous |
Pennsylvanian |
290-320 |
|
|
Mississippian |
320-355 |
||
|
Devonian |
355-410 |
||
|
Silurian |
410-438 |
||
|
Ordovician |
438-510 |
||
|
Cambrian |
510-570 |
||
|
Precambrian |
570-4,600 |
||
The Problem With Dating
Gish argues, “It should be realized that there is no direct method for determining the age of any rock. While very accurate methods are available for determining the present ratios of uranium-lead, thorium-lead, potassium-argon, and other isotope ratios in mineral-bearing rocks, there is, of course, no direct method for estimating the initial ratios of these isotopes in the rocks when the rocks were first formed. Radiochronologists must resort to indirect methods which involve certain basic assumptions. Not only is there no way to verify the validity of these assumptions, but inherent in these assumptions are factors that assure that the ages so derived, whether accurate or not, will always range in the millions to billions of years.”[7]
The radiometric methodology seems to lend credence to the evolutionary theory, but even this methodology is based on unverifiable assumptions, and does not really in practice provide a standard.
Gish notes: “It is often suggested, however, that radiometric dating methods are used to date fossils, a method entirely independent of any supposed stratigraphic correlation or evolutionary stories. Derek Ager, Professor of Geology at University College, Swansea, Wales, has reacted angrily to such claims. He says,
‘My frustrations as a geologist were brought to boiling point by David Challinor’s article on natural history museums (New Scientist, 29 September, 1983, p. 959) and in particular by his remark that “Originally, paleontologists dated fossils by identifying the geological strata in which they were found. Today the age of a fossil is determined by measuring the decay of radioactive carbon or by means of the decay of their radioactive potassium in argon” … Ever since William Smith at the beginning of the 19th century, fossils have been and still are the best and most accurate method of dating and correlating the rocks in which they occur. … As for having all the credit passed to the physicists and the measurement of isotopic decay, they make my blood boil! Certainly such studies give dates in terms of millions of years, with huge margins of errors. … I can think of no cases of radioactive decay being used to date fossils.’[8]
Thus it seems that fossils are used to date rocks, not radiometric dating methods. That seems to bring us back into a circle, for how do we date fossils? In the final analysis, all appears to rest on an assumed evolutionary sequence.”[9]
Molecular data
“The molecular data, however, muddies the water as badly as do the fossils. For example, Weishampel, Dodson, and Osmolska state:
Molecular data on tetrapod phylogeny are equivocal regarding the relationship of birds and crocodiles. Some analyses pair these two groups … but many tend to link birds and mammals more closely … however, other protein sequence analyses give every other imaginable pairing of tetrapod groups, and their significance is debatable …[10]
It seems obvious that if protein sequence analyses gives every imaginable pairing of tetrapod groups, such data are useless in determining the relationships of these groups.”[11]
Lack of transitional forms:
One of the strongest cases against evolution is the total absence of transitional forms. Darwin himself recognized this:
“The number of intermediate varieties, which have formerly existed [must] truly be enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely-graduated organic chain; and this, perhaps, is the most obvious and serious objection which can be urged against the theory [of evolution].” Darwin, The Origin of Species, p. 323.[12]
Walt Brown also provides the following quote:
“Well, we are now about 120 years after Darwin and the knowledge of the fossil record has been greatly expanded. We now have a quarter of a million fossil species but the situation hasn’t changed much. The record of evolution is still surprisingly jerky and, ironically, we have even fewer examples of evolutionary transition than we had in Darwin’s time. By this I mean that some of the classic cases of darwinian change in the fossil record, such as the evolution of the horse in North America, have had to be discarded or modified as a result of more detailed information - what appeared to be a nice simple progression when relatively few data were available now appears to be much more complex and much less gradualistic. So Darwin’s problem has not been alleviated in the last 120 years and we still have a record which does show change but one that can hardly be looked upon as the most reasonable consequence of natural selection.” David M. Raup, ‘Conflicts Between Darwin and Paleontology,” Field Museum of Natural History Bulletin, Vol. 50, No. 1, January 1979, p. 25.
Case Study: From Microorganisms To Fish
Duane Gish notes this problem as follows: “We will now describe the many recent publications that discuss the pervasive, perplexing, and persistent problem for evolutionary theory due to the explosive appearance of a vast array of complex invertebrates in the fossil record with a total absence of ancestors and no trace of transitional forms between the various kinds of invertebrates. Richard Dawkins, the British biologist and evolutionist, states:”[13]
… the Cambrian strata of rocks, vintage about 600 million years [evolutionists are now dating the beginning of the Cambrian at about 530 million years], are the oldest in which we find most of the major invertebrate groups. And we find many of them already in an advanced state of evolution, the very first time they appear. It is as though they were just planted there, without any evolutionary history. Needless to say, this appearance of sudden planting has delighted creationists.[14]
And Gish also quotes Douglas Futuyma, in his book on evolutionary biology as follows:
“It is considered likely that all the animal phyla became distinct before or during the Cambrian, for they all appear fully formed, without intermediates connecting one form to another.”[15]
This lack of transitional forms is a major theme of the fossil record as can be seen in case after case.
One of the major tenets of Darwinian gradualism is that evolution is slow and takes a lot of time (due to the extreme rarity of beneficial radiation induced mutations). However, the lack of fossil evidence forces them to “squeeze” major sections of evolution into increasing smaller time periods in order to explain the lack of fossil evidence. This is the case for the evolution from invertebrates to vertebrates. Gish notes: “As more and more discoveries are made, evolutionists are getting squeezed more and more. They used to date the … Cambrian period … duration … [at] about 80 million years. Now they are assigning a … time span which they believe may encompass no more than ten million years and most likely only five million years.”[16]
The “sudden” diversification of life in the Cambrian layer is baffling to evolutionists. Gish quotes a Swedish paleontologist:
“Baffling (and embarrassing) to Darwin, this event still dazzles us … The animal phyla emerged out of the Precambrian mists with most of the attributes of the modern descendants.”[17]
Gish notes: “Yes, indeed, this sudden appearance of complex invertebrates ‘out of the Precambrian mist’ without a trace of ancestors or transitional forms is still baffling and embarrassing to evolutionists today, just as it was to Darwin, because 135 years after Darwin evolutionists are no nearer to a solution of the ‘mystery’ than was Darwin.”[18]
While evolutionists cling to the concept, some are frank about the lack of evidence. Gish quotes a paleontologist of the University of Cambridge who has studied Cambrian fossils extensively as follows:
“The ‘Cambrian explosion’ is a real evolutionary event, but its origins are obscure. At least 20 hypotheses have been proposed, and although arguments linking diversification to oxygen levels, predation, faunal provinciality, and ocean chemistry all attract support, it is the case that ‘The emergence of Metazoa remains the salient mystery in the history of life.’”[19]
In other words, evolutionists cling to a theory that cannot predict or explain the evidence. Gish notes that one of the most thorough discussions of the “Cambrian explosion” is found in “Origin and Early Radiation of the Metazoa”, by paleontologists Philip Signor and Jere Lipps. They report:
“… there is little evidence that the capacity to form skeletons was acquired gradually or over a prolonged period … A wide variety of skeleton types and most of the major marine invertebrate clades appear suddenly in the fossil record. … The ecological diversification of animals is equally dramatic.”[20]
The admitted mechanism for evolution requires gradualism (a long period of time), and yet the evidence is non-existent, since the invertebrates appear “suddenly” in the record.
Gish notes four major arguments used by evolutionists to explain the fossil record:
Hidden evolution. This argument requires a long period of evolution that is “hidden” (or missing from the fossil record), but must have existed since those creatures that do abruptly appear are very complex and very diverse. “It is pointed out, for example, that fully developed trilobites appear throughout the world, no primitive intermediate forms have ever been discovered …” Therefore, it is assumed that since other cases have the same “problem”, that it is not without precedent. Gish notes “This scenario is based on nothing more, of course, than the assumption of evolution.”[21]
Skeletons. Gish notes that many evolutionists have held that the lack of transitional forms is because these forms were all soft-bodied and that soft-bodied animals generally produce “few fossils.” Gish notes: “The alleged discoveries of fossils of microscopic, single-celled, soft-bodied bacteria and algae have appeared frequently in scientific literature during the past few decades. … Furthermore, large numbers of fossils of soft-bodied creatures, such as jellyfish and worms, have been found, many in a remarkable state of preservation.”[22]
Atmospheric Oxygen. Some have suggested that the Cambrian explosion of complex vertebrates was made possible by the accumulation of free oxygen in the atmosphere. If this were true, then one would expect previous layers (periods) to have demonstrably less oxygen levels. Gish notes: “Most geological research in recent years has established beyond little doubt that the earth has always had an oxygenated atmosphere, certainly since sedimentary rocks first began to form.”[23] He quotes Signor and Lipps: “… there is no evidence that oxygen was limiting in the late Proterozoic.”[24]
Ocean Chemistry. Some have argued that certain aspects of the chemistry of the oceans prior to the origin of complex invertebrates inhibited their formation. Gish notes: “But as Signor and Lipps point out, skeletons of calcium carbonate, calcium phosphate, biogenic silica, and agglutinated skeletons all appeared simultaneously in a great variety of invertebrates.”[25] In other words, if this explanation is true, then the chemistry change was a complex one that allowed all skeletal forms to progress simultaneously -- not a likely scenario in Darwinian gradualism.
None of these are generally held, and the lack of any reasonably valid explanation is a deep blow to the case of evolution. However, all of the evidence is just what creation demands -- a “sudden” appearance of fully developed forms, just as God created them.
Case study: Origin of Reptiles
Reptiles, birds, and mammals are classified as Amniota and therefore referred to as amniotes. Although evolutionists are hard pressed to “find” real evidence for transitional forms, they do produce arguments for some forms. The evolution argument calls for reptiles to be evolved from amphibians. Some have attempted to find creatures which have common features with both amphibians and reptiles, and use them as transitional forms. One such attempt is the Seymouria. Gish quotes Colbert and Morales as follows:
“The mixture of amphibian and reptilian characters seen in Seymouria is indicative of the gradual transition that took place between the two classes during the evolution of the vertebrates.”[26]
Gish Counters: “Those creatures suggested as the earliest known reptiles, Hylonomus and Paleothyris, are found in Lower Pennsylvanian rocks (about 330-315 million years before present on the evolutionary time scale) and the Middle Pennsylvanian rocks (about 310-315 million years), respectively. Fossils of Seymouria are found in Lower Permian rocks, dated at about 280 million years, or at least 25-35 million years too late to be ancestral to reptiles.”[27]
“Diadectes is often cited as being very close to the dividing line between amphibians and reptiles. It is also a most improbable candidate, however. It is about 30 million years too late on the evolutionary time scale to be ancestral to reptiles …“[28]
Gish quotes from Carroll -- “The early amniotes are sufficiently distinct from all Paleozoic amphibians that their specific ancestry has not been established (p. 198).”[29] And Gish adds, “The last statement just quoted establishes beyond doubt that transitional forms between amphibians and reptiles do not exist. If they had been found, there would be no doubt concerning which amphibian gave rise to reptiles.”[30]
Case Study: Archaeopteryx (Birds & Reptiles / Dinosaurs)
Archaeopteryx -- from “Archae” (ancient) and “pteryx” (wing).
“It has long been accepted that Archaeopteryx was a transitional form between birds and reptiles, and that it is the earliest known bird. Lately, scientists have realized that it bears even more resemblance to its ancestors, the Maniraptora, than to modern birds; providing a strong phylogenetic link between the two groups. It is one of the most important fossils ever discovered.”[31]
“In a book entitled "Taking Wing" by the American anthropologist PAT SHIPMAN (4), dealing with current debate on the origin and early evolution of birds, the author writes about the seven Archaeopteryx specimens: “[32]
"They include the most beautiful fossil in the world; the most famous fossil in the world; the jewel in the crown of the most beautiful museum in Europe; the proof of evolutionary theory; the evidence of the origin of birds; the secret of bird flight. How can seven scant specimens be so significant? How can it be otherwise?" (underlining mine, D.A.D.)
Evolutionists have maintained that birds have evolved from reptiles. However, some argue that birds are evolved from dinosaurs, with some saying that they are most closely related to crocodiles. The Archaeopteryx is at the center of the controversy. Gish notes “An International Archaeopteryx Conference was held in Eichstatt, Bavaria, September 11-15, 1984, during which advocates of the above ideas put forth their arguments.[33] Especially interesting was the fact that those attending the conference felt it necessary to adopt a communiqué ‘expressing the unanimous belief of all participants in the evolutionary origin and significance of Archaeopteryx in order to forestall possible misuse by creationists of apparent discord among scientists.’[34] In other words, these evolutionists unanimously agree that evolution is a fact even though they cannot agree on the ancestry of birds, and more particularly on the origin of Archaeopteryx.”[35]
Gish adds, “Ostrom and others have claimed that birds had evolved via feathered ground-dwelling (cursorial) predators that had arisen from coelurosaurian dinosaurs. The claim was made that Archaeopteryx had claws similar to those of ground-dwelling birds rather than similar to those of perching, tree-dwelling birds. D. W. Yalden, however, demonstrated that the claws of the manus (forelimb) of Archaeopteryx were nearly identical to those of tree climbers.[36] … Alan Feduccia … established that the arc of the claws of _Archaeopteryx was comparable to that of tree-dwelling (perching) birds. His analysis established that Archaeopteryx was not a cursorial predator but was a perching bird. Feduccia’s conclusion was that:
‘Archaeopteryx probably cannot tell us much about the early origins of feathers and flight in true protobirds because Archaeopteryx was, in a modern sense, a bird.’[37]”
Gish notes, “Larry D. Martin, an ornithologist and professor of systematics and ecology at the University of Kansa and head of the vertebrate paleontology division in the university’s Museum of Natural History, accepts a modified version of the pseudosuchian ancestry of birds that makes birds a sister group to crocodilians. In a newspaper article originally published in Newsday he states:
‘The theory linking dinosaurs to birds is a pleasant fantasy that some scientists like because it provides a direct entry into a past we otherwise can only guess about. But unless more convincing evidence is uncovered, we must reject it and move on to the next better idea.’[38]
Gish states, “Some evolutionists insist that since Archaeopteryx had characteristics of both reptiles and bird, it does represent an intermediate between reptiles and birds, but whatever features it had were complete, not part-way or transitional.”[39]
Gish states, “It is interesting to note the comment of Stephen Jay Gould of Harvard University and Niles Eldredge of the American Museum of Natural History, both ardent anti-creationists. With reference to Archaeopteryx, they state that:
‘At the higher level of evolutionary transition between morphological designs, gradualism has always been in trouble, though it remains the “offical” position of most western evolutionists. Smooth intermediates between Baupläne [a German word meaning basic morphological designs or basically different types of creatures] are almost impossible to construct, even in thought experiments; there is certainly no evidence for them in the fossil record (curious mosaics like Archaeopteryx do not count).’[40]”
Gish quotes Swinton, an evolutionist and an expert on birds:
“The origin of birds is largely a matter of deduction. There is no fossil evidence of the stages through which the remarkable change from reptile to bird was achieved.”[41]
Gish also quotes Romer:
“This Jurassic bird (Archaeopteryx) stands in splendid isolation; we know no more of its presumed thecodont ancestry nor of its relation to later “proper” birds than before.”[42]
In addition to these difficulties, there is the problem of finding birds that are older than Archaeopteryx. Walt Brown notes: “Significantly, two modern birds have recently been found in rock strata dated by evolutionists as much older than Archaeopteryx.[43] Therefore, Archaeopteryx could not be ancestral to modern birds.”[44]
Case Study: The Horse
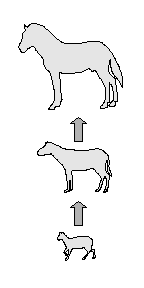
“A case in point is the evolution of the modern horse, which is believed to be a descendant of a much smaller ancestor named Hyracotherium, which browsed in the woods of the Eocene epoch about 40 million years ago. In comparison to the ancestor, not only are modern horses (genus Equus) larger, but the number of toes has been reduced from four on each foot to one, and the teeth have become modified for grazing rather than browsing. By selecting certain species from the available fossils, it is possible to arrange a succession of animals intermediate between Hyracotherium and modern horses that shows trends toward increased size, reduced number of toes, and grazing teeth. We might interpret this series of fossils as an unbranched lineage leading directly from Hyracotherium to modern horses through a continuum of intermediate stages. If we include all fossil horses known today, however, the illusion of coherent, progressive evolution leading directly to modern horses vanishes. The transition occurs in steps rather than in a smooth gradation of forms; each species appears and disappears in the fossil record without changing noticeably in the interim.”[45] (underlining mine -- D.A.D.) This reference still teaches the evolution of the horse with slight modification from the original, but this admission that an arbitrary illusion of evolution can be created by placing animal in sequence from small to large is damaging. A careful observer might well ask the question “How is the new ‘proposed’ sequence any more convincing that the first ‘illusion of evolution’ ”?
The reader may further ask the question, “How is the first proposed ‘illusion’ different from any other proposed evolutionary sequence?” That is a very good question. Gish notes: “In November of 1980, 150 evolutionists met for four days at the Field Museum of Natural History in Chicago to attack or defend the gradualistic evolutionary theory, or the new-Darwinian theory of evolution. … Those attacking the new-Darwinian mechanism of evolution maintained there is little or no evidence of gradualism in the fossil record. Naturally the story of the alleged evolution of the horse was discussed. Boyce Rensberger, in his report of the meeting, stated that:
‘The popularly told example of horse evolution, suggesting a gradual sequence of changes from four-toed fox-size creatures living nearly 50 million years ago to today’s much larger one-toed horse, has long been known to be wrong. Instead of gradual change, fossils of each intermediate species appear fully distinct, persist unchanged, and then become extinct. Transitional forms are unknown.[46]’
In other words, the alleged horse story is yet another in a long line of acknowledged inabilities of Darwinian evolution to predict or explain the fossil evidence. The persistent lack of any transitional forms is a clear indictment of evolution.
What About Plants?
Gish records, “The following remarks of E. J. H. Corner of the Cambridge University botany school were refreshingly candid:”[47]
“Much evidence can be adduced in favor of the theory of evolution -- from biology, biogeography, and paleontoloty, but I still think that to the unprejudiced, the fossil record of plants is in favor of special creation.”[48]
Punctuated Equilibrium
The profound lack of fossil evidence has prompted many, including Ernst Mayr who is sometimes described as having one of the “greatest influences on evolutionary biology since Darwin”[49], to embrace what is commonly termed “punctuated equilibrium”. Ernst Mayr is quoted as saying “A mistake people make is thinking that if something evolves very rapidly, it is no longer Darwinian gradual evolution. But as long as the evolution occurs at the populational level and not at the level of individuals, then it is gradual evolution, occurring over many generations. Some paleontologists call the origin of a species relatively sudden if it takes place during 1% of the total lifespan of the species. But that 1% is 50,000 years or 100,000 years in a species living 5 or 10 million years. Hardly anyone else would call that sudden”
“In the most common occurrence of speciation, according to punctuated equilibrium, small, isolated populations evolve rapidly, so that speciation takes place over about ten thousand to a million years. This period of time is a geological instant, but it still allows plenty of time for gradual change at a fast rate to produce a new species.
Punctuated equilibrium explains how large, stable populations can produce new species: the large population itself doesn't change, but small isolated "pocket" populations might, resulting in branching rather than linear species histories. It also explains the relative scarcity of transitional forms, particularly between species (rather than between larger groups), in the fossil record. If transitional forms only exist for a few thousand years, often in a small geographical location different from its later range, then the odds are against fossils being formed, found, and described.”[50]
Problems with the geological sequence:
Rapid Burial
Walt Brown notes: “Fossils all over the world show evidences of rapid burial. Many fossils, such as fossilized jellyfish, show by the details of their soft, fleshy portions that they were buried rapidly, before they could decay. Many other animals, buried in mass graves and in twisted and contorted positions, suggest violent and rapid burials over large areas. These observations, together with the occurrence of compressed fossils and fossils that cut across two or more layers of sedimentary rock, are strong evidence that the sediments encasing these fossils were deposited rapidly - not over hundreds of millions of years. Furthermore, almost all sediments were sorted by water. The worldwide fossil record is, therefore, evidence of the rapid death and burial of animal and plant life by a worldwide, catastrophic flood. The fossil record is not evidence of slow change.”[51]
Gradualism
The earth’s geological record shows a lot of evidence for a world-wide catastrophe which would lay down tremendous volumes of material very rapidly and vast changes in the environment. The large numbers of Mammoths that have been discovered testify to this. These are largely found inside the Arctic Circle and show evidence of being rapidly buried in ice. The flesh is so well preserved that dogs have been known to eat the flesh of thawing Mammoths. For this situation to occur, the Mammoth must have been buried and frozen at an astonishingly rapid rate to prevent decomposition. The primary cause of death is usually attributed to suffocation.
Walt Brown records the following evidence with regard to Mammoths:[52]
“Sanderson describes another strange aspect of Bere˗zovka.
Much of the head, which was sticking out of the bank, had been eaten down to the bone by local wolves and other animals, but most of the rest was perfect. Most important, however, was that the lips, the lining of the mouth and the tongue were preserved. Upon the last, as well as between the teeth, were portions of the animal’s last meal, which for some almost incomprehensible reason it had not had time to swallow The meal proved to have been composed of delicate sedges and grasses
“Another account states that the mammoth’s ‘mouth was filled with grass, which had been cropped, but not chewed and swallowed.’ The grass froze so rapidly that it still had ‘the imprint of the animal’s molars.’ Hapgood’s translation of a Russian report mentions eight well-preserved bean pods and five beans found in its mouth.”
“Twenty-four pounds of undigested vegetation were removed from the Berezovka mammoth and analyzed by the Russian scientist, V. N. Sukachev. He identified more than forty different species of plants: herbs, grasses, mosses, shrubs, and tree leaves. Many no longer grow that far north; others grow both in Siberia and Mexico. Dillow draws several conclusions from these remains:
• The presence of so many varieties [of plants] that generally grow much to the south indicates that the climate of the region was milder than that of today
• The discovery of the ripe fruits of sedges, grasses, and other plants suggests that the mammoth died during the second half of July or the beginning of August.
• The mammoth must have been overwhelmed suddenly with a rapid deep freeze and instant death. The sudden death is proved by the unchewed bean pods still containing the beans that were found between its teeth, and the deep freeze is suggested by the well-preserved state of the stomach contents and the presence of edible meat [for wolves and dogs]”[53]
The evidence for catastrophic change denies one of the foundation premises of evolutionary dating mechanisms which “assume” uniformity.
Statistics
The principle mechanism of evolutionary change is that of mutation. Mutations are very rare, statistically speaking, and this is usually acknowledged in the long time spans given for evolution. Many have argued that statistically speaking the “chances” of evolution producing man from molecules is virtually nil. Even evolutionists admit that the likelihood of a “good” mutation is highly unlikely. Neil Campbell in his textbook on Biology says:
“However, mutation by itself does not have much quantitative effect on a large population in a single generation. This is because a mutation at any given gene locus is a very rare event; although mutation rates vary, depending on the species and the gene locus, rates of one mutation per locus per 105 to 106 gametes are typical. If an allele has a frequency of 0.50 in the gene pool and mutates to another allele at a rate of 10-5 mutations per generation, it would take 2000 generations to reduce the frequency of the original allele from 0.50 to 0.49. The gene pool would be affected even less if the mutation were reversible, as most are.”[54]
In addition, with regard to “good” mutations he says:
“Organisms are the refined products of thousands of generations of past selection, and a random change is not likely to improve the genome any more than firing a gunshot blindly through the hood of a car is likely to improve engine performance.”[55]
Actually, this is a quite fitting analogy, and I don’t know anyone who would logically argue that firing a gunshot blindly through the hood of a car would improve engine performance -- no matter how many bullets were used!
The Problem Compounded
The problems of statistics in evolution is compounded when the evidence demands that multiple species all mutate at the same time. Duane Gish quotes Romer concerning archosaur evolution as follows:
“Many similarities in structural features among end forms of different archosaurian lines have not been inherited as such from a common ancestor but have been independently acquired by members of the different groups. This however, does no debar such characters from consideration as indications of relationship. Study of fossil forms increasingly indicates that there has been an enormous amount of parallelism in evolution; but this study also appears to demonstrate that close parallelism occurs only in closely related forms.”[56]
This “parallel evolution”, which seems to be demanded by the evidence (when each of the descendant lines has the same feature which the supposed parent did not) as viewed by evolutionists occurs in “enormous” amounts. Now if “good” mutations are extremely rare, such that large numbers of generations are required to influence the population, and the statistical odds are very, very small, then what are the odds that the “same” “good” mutation occurs during the same time period to “many” species? If it happened just twice, the odds would be very small, but to occur “many” times in parallel, it is unbelievable. An unbelievable chance occurrence happening many time in parallel -- blind faith is called for here!
More Parallelism
With regard to the evolution of mammals, Gish quotes Kemp as follows:
“It was noted that the fossil record supports the view that evolution towards mammalian levels of homeostasis involved practically all aspects of the organism simultaneously. No single structure or function could evolve very far without being accompanied by appropriate changes in all the other features.”[57]
In this case extremely rare mutations are required in not one area alone, but in parallel since each of the features being evolved could not continue without the parallel development of others.
Conclusion
The compounded parallelism of evolution requires rare events to happen during the same time interval in multiple species, and during the same time interval within species, and furthermore this whole scenario is compounded by having to occur in increasingly short intervals of time. The time element is shortened of necessity to explain the lack of transitional forms (i.e. “punctuated equilibrium”). Roll it all together, and the end result is that statistically speaking, evolution is impossible.